
Ocean temperature and chemistry changes strongly influence the well-being of organisms and the composition of marine food webs. Since the preindustrial era, the oceans have absorbed at least one fourth of the carbon dioxide (CO2) that humans have put into the atmosphere, resulting in an increase in ocean acidity. The amount of oxygen dissolved in ocean water has changed as well, driven by warming and changes in circulation patterns. These effects are further compounded in coastal regions, where marine ecosystems face additional human pressures such as pollution, land use changes, and overfishing.
Long-term, seasonally resolved time series observations in the ocean have provided unprecedented insights into how ocean biogeochemistry and ecosystems are changing on a range of temporal scales. We now know that these observations must be sustained for a minimum of several decades to statistically distinguish natural climate variability from changes associated with human activities [e.g., Henson et al., 2016]. Here we present a scientific vision for the future of shipboard biogeochemical time series in light of new scientific findings and recent technological advances.
Ocean time series stations have documented major changes in the chemistry of ocean waters, in the functioning of ecosystems, and in oceanic carbon sequestration. These stations have increasingly served as test beds for the development of new sensors and methodologies. They provide valuable seagoing opportunities and hands-on training for the next generation of ocean scientists. Ocean time series efforts are also a forum for collaboration between nations, building scientific capacity. Efforts to establish a global network of observatories are underway: If these efforts are sustained into the future, they will allow us to monitor how global marine ecosystems are responding to a changing climate.
Balancing the Carbon Budget for Marine Ecosystems
Long-term biogeochemical and ecological time series observations have yielded fundamental insights into processes controlling carbon exchange between the air and sea, rates of carbon transformation throughout the marine food web, and fluxes of carbon into the ocean’s interior.
For example, in the subtropical ocean gyres, the annual net community production (NCP) and the air-sea exchange of CO2 dominate the flow of carbon near the ocean’s surface [e.g., Quay and Stutsman, 2003]. NCP measures the difference between how much organic carbon autotrophs (organisms at the base of the marine food web) make through photosynthesis (primary production) and how much carbon heterotrophs (microbes and animals that consume phytoplankton and their waste products) release through respiration (primary production minus respiration). NCP reflects the ocean’s capacity to biologically sequester atmospheric CO2 for periods of months to millennia by exporting organic carbon to the deep sea.

Quantifying NCP with high confidence and reproducibility represents a key challenge for the oceanographic community. Over the past 2 decades, ocean time series programs have provided seasonal observations for quantifying NCP, including a new suite of ocean productivity measurements that supplement shipboard observations with autonomous and remote sensing platforms and sensors [e.g., Church et al., 2013]. Systematic comparison of these approaches has proven invaluable in identifying pathways that regulate carbon fluxes and testing assumptions and uncertainties underlying productivity measurements and models.
Several ocean time series sites show marked imbalances between NCP and annual export flux or export production (EP). The EP is a measure of particulate organic matter that sinks through the water and is eventually sequestered in the deep ocean. EP is measured by capturing organic particles in sediment traps or estimated from satellite-derived productivity models [e.g., Emerson, 2014].
At the Bermuda Atlantic Time-series Study (BATS) site in the northwest Sargasso Sea, estimated NCP is nearly 3 times the annual EP. Remarkably, similar NCP and NCP/EP imbalances occur in the North Pacific subtropical gyre at the Hawaii Ocean Time-series (HOT) site and the subarctic northeast Pacific at Ocean Station Papa, and the largest imbalance is observed in the eastern subtropical North Atlantic at the European Station for Time series in the Ocean at the Canary Islands (ESTOC) [e.g., Helmke et al., 2010].
Ongoing research at time series stations has shown that imbalances between NCP and EP may be partially reconciled by taking into account the physical mixing of suspended organic particles and dissolved organic matter. Other factors include zooplankton fecal pellets’ ability to sink rapidly and thus transport carbon away from the surface layer of the ocean, coupled with active zooplankton migration, variations in respiration with depth, and departures from well-established [e.g., Redfield, 1934] marine plankton and organic matter carbon to nitrogen to phosphorus ratios.
Chemistry Changes and Marine Ecosystems
The oceans have absorbed 25%–30% of the anthropogenic CO2 emitted since the preindustrial era [Sabine et al., 2004]. Recent analyses of ocean CO2 across multiple independent ocean time series sites show increasing acidity across ocean basins over the past 2–3 decades [e.g., Bates et al., 2014; Tanhua et al., 2015]. This widespread change in the ocean CO2 system is a direct consequence of ocean uptake of CO2 (Figure 1). These studies demonstrate the global extent of this phenomenon and underscore the importance of high-resolution data sets to remove seasonality and elucidate longer-term trends.
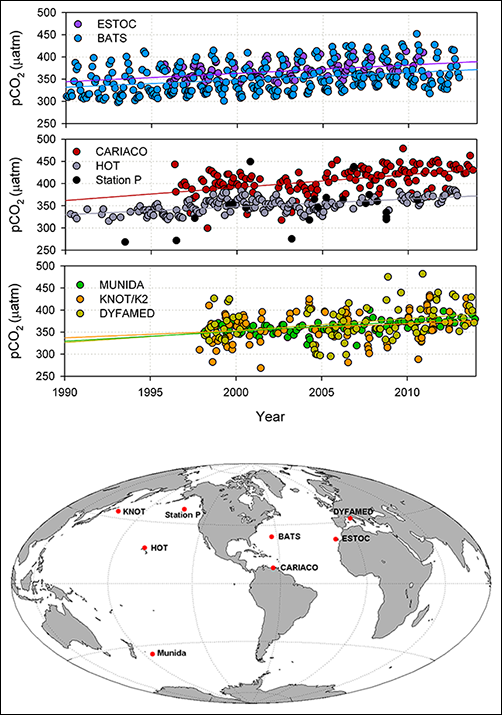
In addition to increasing acidity, the oceans have undergone warming, increased stratification, and circulation changes that have reduced dissolved oxygen levels, particularly in coastal regions like the southern California Current System. The California Cooperative Oceanic Fisheries Investigations (CalCOFI) ocean time series has documented a decline in dissolved oxygen levels and a shoaling of the oxygen minimum layer (a relocation of a region of the water column with very low dissolved oxygen levels to shallower waters) over the past 2–3 decades. These changes are likely to affect organisms living in the water column and on the seafloor [Bograd et al., 2008].
Two decades of observations (1996–2016) from the Carbon Retention in a Colored Ocean (CARIACO) ocean time series in the Cariaco Basin off the northern coast of Venezuela have documented similarly important climate and ecosystem changes in the tropical Atlantic. In 2004, the phytoplankton community in this region underwent a marked shift from mostly diatoms to much smaller phytoplankton, accompanied by increased phytoplankton diversity and zooplankton biomass and declining primary productivity. These changes were accompanied by decreases in chlorophyll a (the type of chlorophyll involved in photosynthesis) and surface nitrate concentrations that reflect major adjustments in the circulation and biogeochemistry of the Atlantic [Montes et al., 2016].
Ocean time series have documented seasonal changes that can affect the entire marine food web.
Ocean time series have also documented changes in the seasonal occurrence (phenology) of organisms at lower trophic levels, which can affect the entire marine food web, including major U.S. fisheries. A recent study reporting data from an ocean time series in New England coastal waters [Hunter-Cevera et al., 2016] showed that phytoplankton blooms now occur earlier in the spring in response to warmer water temperatures. Such climate-driven shifts in bloom phenology affect the function of marine food webs and, ultimately, alter the ocean’s capacity to provide food and sequester carbon. A recent study using 57 years’ worth of data from the CalCOFI time series [Asch, 2015] revealed shifts in the phenology of larval fishes tied to earlier surface ocean warming in the California Current ecosystem, a productive commercial fishery.
Ocean time series observations of chemical and biological parameters in marine ecosystems can also help inform the management of marine resources. For example, a recent study from the northern California Current [McKibben et al., 2017] documented an increased incidence of shellfish containing domoic acid (a potent neurotoxin produced by some phytoplankton) associated with warm sea surface temperature anomalies during the upwelling season over the past 20 years. This warm water could be tied to the warm phases of the Pacific Decadal Oscillation (PDO) and the El Niño–Southern Oscillation (ENSO). A risk assessment model based on this connection of climatic and local events can predict domoic acid outbreaks along the U.S. West Coast and other eastern boundary current systems worldwide.
Test Beds for New Technology

Ocean time series programs provide routine access to the sea through the regular availability of research vessels and high-quality data sets needed to support development of new methodologies, technologies, and calibration and validation activities (e.g., satellite algorithm development, paleoproxy development, model-based prediction). Time series data also serve as important benchmarks with which Earth system model outputs are compared.
A symbiosis exists between shipboard time series and measurements from autonomous platforms such as moorings, profiling floats, and gliders. Autonomous sensors (e.g., gliders deployed at and around a time series site) expand the observational footprint of ocean time series by capturing variability on shorter time scales and by enabling broader spatial coverage. For example, sensors deployed on moorings and profiling floats have provided insight into the episodic physics that contribute significantly to biogeochemical variability at the time series locations. High-frequency measurements have repeatedly demonstrated that monthly shipboard measurements miss episodic events that affect seasonal and interannual biogeochemical variability. In turn, in situ measurements from shipboard time series provide critical information for calibrating and interpreting data from autonomous sensors.
A Global Time Series Network: A Whole Much Greater Than the Sum of Its Parts
In addition to monitoring changes in marine ecosystems with unprecedented resolution, time series programs have provided unique and cost-effective opportunities to support the scientific goals of countless ancillary projects. This is evident in the rich body of literature that has emerged from ocean time series data (e.g., publication lists from CARIACO, HOT, and BATS).
Monitoring ocean change requires a sustained, globally distributed network of observatories that integrates shipboard, autonomous, and remote sensing platforms.
U.S.-funded time series programs such as BATS, HOT, CalCOFI, and CARIACO have hosted hundreds of ancillary projects, from small student projects to large research collaborations with multiple primary investigators. They have also yielded preliminary data that investigators can use to develop full partnership programs such as the Center for Microbial Oceanography: Research and Education, the National Science Foundation’s Microbial Observatory program, glider and mooring programs, the Simons Foundation collaborations with HOT and BATS, and the Nippon Foundation–Partnership for Observation of the Global Oceans (NF-POGO) Centre of Excellence in Observational Oceanography.
Monitoring ocean change requires a sustained, globally distributed network of observatories that integrates shipboard, autonomous, and remote sensing platforms. The ultimate goal of such a network is to obtain a broader spatial perspective of how our marine ecosystems are changing through time, which is possible only if data from various time series stations remain comparable through standardized methodological approaches and through a willingness to share data openly. Several initiatives spanning ocean time series have taken place in recent years to work toward the goals of improved data intercomparability and accessibility.
For example, in November 2012, the U.S.-led Ocean Carbon and Biogeochemistry (OCB) Program and the International Ocean Carbon Coordination Project (IOCCP) co-organized an International Time-series Methods Workshop at the Bermuda Institute of Ocean Sciences [Benway et al., 2013]. This workshop convened representatives from global marine biogeochemical time series sites to review current shipboard biogeochemical time series sampling and analytical protocols. The outcome was a global compilation of measurements and methods used at shipboard biogeochemical time series sites, a best practices guide [Lorenzoni and Benway, 2013], and a permanent website and email list to support a global shipboard biogeochemical time series network.

Investigators in the International Group for Marine Ecological Time Series, another ongoing international collaboration of shipboard ocean biogeochemical time series, have integrated a suite of in situ biogeochemical variables from more than 300 globally distributed time series stations with satellite-derived information about trends in ocean temperature and chlorophyll. Their objectives are to identify holistic changes within different ocean regions over the past 1 to 3 decades, to explore plausible connections at a global level, and to highlight regions of the ocean that are undergoing especially large biogeochemical and ecological changes [O’Brien et al., 2017].
Efforts are also underway to develop robust autonomous observing networks and facilitate more widespread use of autonomous sensors. OceanSITES is an international observing network consisting of long-term, open-ocean reference stations measuring dozens of variables and monitoring the full depth of the ocean from air-sea interactions down to the seafloor, including observations of meteorology, physical oceanography, biogeochemistry, and geophysics. Biogeochemical Argo is another emerging autonomous ocean-observing initiative focused on developing a global network of biogeochemical sensors on Argo profiling floats.
Recent training efforts have focused on establishing community-accepted best practices to guide deployment of autonomous biogeochemical sensors and develop a common set of standards to improve the quality of the data these sensors collect. These efforts are aimed at increasing sensor development and deployment and enhancing the oceanographic community’s proficiency in the use of biogeochemical sensors (e.g., oxygen, pH, CO2 partial pressure, and nitrate).
Our Vision for the Future
Collaborative efforts across the global network of ocean time series are driven by scientific challenges and questions. Many of these challenges and questions emerge from societally relevant problems, or they eventually have direct applications. Science-driven synthesis efforts across time series sites maximize the scientific return on investment in these observatories. They make the best use of existing observational capacities, identify knowledge gaps, and guide plans for future observations to address these gaps.
Working effectively and routinely across time series sites will require coordination between international funding agencies, a culture of mutual trust and respect between generators and users of time series data, and effective mechanisms to incentivize data sharing (e.g., data publications with digital object identifiers, or DOIs) and data management and interoperability across ocean data centers.
Ultimately, this global network is built upon the collaborations forged among scientists who generate and work with ocean time series data.
Underlying any successful effort to build and sustain ocean-observing networks is the need for long-term support from funding agencies, as well as initiative and leadership from within the oceanographic community to bring the international community of scientists together around a common goal.
Ongoing initiatives such as the NF-POGO Alumni Network for Oceans represent important opportunities to build capacity and support collaboration, education, and communication across the international ocean sciences community. Strengthening ties to other established ocean-observing networks such as the Ocean Observatories Initiative, OceanSITES, Biogeochemical Argo, and the Long Term Ecological Research Network is an important near-term priority for an international time series network.
Ultimately, this global network is built upon the collaborations forged among scientists who generate and work with ocean time series data. We place a high priority on strengthening connections between multidisciplinary time series programs and modelers who use the observations.
We advocate for the time series community to continue to invest in activities, as described above, that bring together time series scientists such as workshops, intercomparison activities (calibration, methodological testing, development and testing of standards and reference materials, interdisciplinary interpretation of data, etc.), training activities, and outreach efforts (e.g., a shared library of educational tools and resources on ocean time series). These activities will facilitate collaboration and coordination among scientists and across sites, paving the way for transformative science and a thriving observational network.
Acknowledgments
This is a contribution of the Ocean Time-series Subcommittee of the OCB program with input from presentations and discussion held at the 24 February 2016 Ocean Sciences Meeting in New Orleans, La., on “The Future of Biogeochemical Ocean Time Series.”
References
Asch, R. (2015), Climate change and decadal shifts in the phenology of larval fishes in the California Current ecosystem, Proc. Natl. Acad. Sci. U. S. A., 112, E4065–E4074, https://doi.org/10.1073/pnas.1421946112.
Bates, N. R., et al. (2014), A time-series view of changing ocean chemistry due to ocean uptake of anthropogenic CO2 and ocean acidification, Oceanography, 27(1), 126–141, https://doi.org/10.5670/oceanog.2014.16.
Benway, H. M., M. Telszewski, and L. Lorenzoni (2013), Improving intercomparability of marine biogeochemical time series, Eos Trans. AGU, 94(17), 160, https://doi.org/10.1002/2013EO170004.
Bograd, S. J., et al. (2008), Oxygen declines and the shoaling of the hypoxic boundary in the California Current, Geophys. Res. Lett., 35, L12607, https://doi.org/10.1029/2008GL034185.
Church, M. J., M. W. Lomas, and F. M. Karger (2013), Sea change: Charting the course for biogeochemical ocean time-series research in a new millennium, Deep Sea Res., Part II, 93, 2–15, https://doi.org/10.1016/j.dsr2.2013.01.035.
Emerson, S. R. (2014), Annual net community production and the biological carbon flux in the ocean, Global Biogeochem. Cycles, 28, 14–28, https://doi.org/10.1002/2013GB004680.
Helmke, P., et al. (2010), Cross-basin differences in particulate organic carbon export and flux attenuation in the subtropical North Atlantic gyre, Deep Sea Res., Part I, 57, 213–227, https://doi.org/10.1016/j.dsr.2009.11.001.
Henson, S. A., C. Beaulieu, and R. Lampitt (2016), Observing climate change trends in ocean biogeochemistry: When and where, Global Change Biol., 22, 1,561–1,571, https://doi.org/10.1111/gcb.13152.
Hunter-Cevera, K. R., et al. (2016), Physiological and ecological drivers of early spring blooms of a coastal phytoplankter, Science, 354, 326–329, https://doi.org/10.1126/science.aaf8536.
Lorenzoni, L., and H. M. Benway (Eds.) (2013), Report of Global Intercomparability in a Changing Ocean: An International Time-Series Methods Workshop, November 28–30, 2012, 61 pp., Ocean Carbon and Biogeochem. Program and Int. Ocean Carbon Coord. Proj., Woods Hole, Mass., https://www.us-ocb.org/wp-content/uploads/sites/43/2017/01/TS_Workshop_report_FINAL.pdf.
McKibben, S. M., et al. (2017), Climate regulation of the neurotoxin domoic acid, Proc. Natl. Acad. Sci. U. S. A., 114, 239–244, https://doi.org/10.1073/pnas.1606798114.
Montes, E., et al. (2016), Decadal variability in the oxygen inventory of North Atlantic subtropical underwater captured by sustained, long-term oceanographic time-series observations, Global Biogeochem. Cycles, 30, 460–478, https://doi.org/10.1002/2015GB005183.
Quay, P. D., and J. Stutsman (2003), Surface layer carbon budget for the subtropical N. Pacific: δ13C constraints at station ALOHA, Deep Sea Res., Part I, 50, 1,045–1,061, https://doi.org/10.1016/S0967-0637(03)00116-X.
O’Brien, T. D., et al. (Eds.) (2017), What are marine ecological time series telling us about the ocean? A status report, IOC Tech. Ser. 129, Intergov. Oceanogr. Comm., Paris.
Redfield, A. C. (1934), On the proportions of organic derivations in sea water and their relation to the composition of plankton, in James Johnstone Memorial Volume, edited by R. J. Daniel, pp. 176–192, Univ. Press of Liverpool, Liverpool, U.K.
Sabine, C. L., et al. (2004), The oceanic sink for anthropogenic CO2, Science, 305, 367–371, https://doi.org/10.1126/science.1097403.
Tanhua, T., et al. (2015), Increasing ocean carbon and ocean acidification, WMO Bull., 64(1), 48–51.
Author Information
Susanne Neuer (email: [email protected]), School of Life Sciences, Arizona State University, Tempe; Heather M. Benway, Department of Marine Chemistry and Geochemistry, Woods Hole Oceanographic Institution, Mass.; Nicholas Bates, Bermuda Institute for Ocean Sciences, St. Georges; Craig A. Carlson, Department of Ecology, Evolution, and Marine Biology, University of California, Santa Barbara; Matthew Church, Flathead Lake Biological Station, University of Montana, Polson; Michael DeGrandpre, Department of Chemistry and Biochemistry, University of Montana, Missoula; John Dunne, Geophysical Fluid Dynamics Laboratory, National Oceanic and Atmospheric Administration, Princeton, N.J.; Ricardo Letelier, College of Earth, Ocean, and Atmospheric Sciences, Oregon State University, Corvallis; Michael Lomas, Bigelow Laboratory for Ocean Sciences, East Boothbay, Maine; Laura Lorenzoni, Institute for Marine Remote Sensing, University of South Florida, St. Petersburg; Frank Muller-Karger, College of Marine Science, University of South Florida, St. Petersburg; Mary Jane Perry, University of Maine, Twisp, Wash.; and Paul Quay, School of Oceanography, College of the Environment, University of Washington, Seattle
Correction, 11 September 2017: The photograph of the CTD sampling rosette and crew in this article was replaced on 11 September with a newer photo depicting the crew members wearing safety gear that complies with current standards.
Citation:
Neuer, S.,Benway, H. M.,Bates, N.,Carlson, C. A.,Church, M.,DeGrandpre, M.,Dunne, J.,Letelier, R.,Lomas, M.,Lorenzoni, L.,Muller-Karger, F.,Perry, M. J., and Quay, P. (2017), Monitoring ocean change in the 21st century, Eos, 98, https://doi.org/10.1029/2017EO080045. Published on 08 September 2017.
Text © 2017. The authors. CC BY-NC-ND 3.0
Except where otherwise noted, images are subject to copyright. Any reuse without express permission from the copyright owner is prohibited.