
Recent changes in global climate patterns have redefined what “normal” weather looks like. A “new normal” Arctic has a longer open-water period in summer and wider extremes of weather variability overall.

Before 2010, for example, sea ice covered more than half of the Beaufort and Chukchi seas year-round. Now these seas have less than 50% sea ice cover, and only for an average of 67 days per year [Wood et al., 2015]. Temperature and wind drive changes in sea ice, but the effects are variable across the Pacific Arctic [Frey et al., 2015].
In an effort to provide an interdisciplinary look at completed and ongoing marine research on the Pacific Arctic, a cross-institutional group of researchers launched the Synthesis of Arctic Research (SOAR) in 2011. SOAR delves into the emerging effects of the new normal on Pacific Arctic marine life.
An Interdisciplinary Synthesis
Specifically, SOAR aims to increase scientific understanding of the relationships among such oceanographic conditions as physics, chemistry, and sea ice; benthic (seafloor) organisms; and species occupying various trophic levels (the position an organism occupies in the food web) in this region.
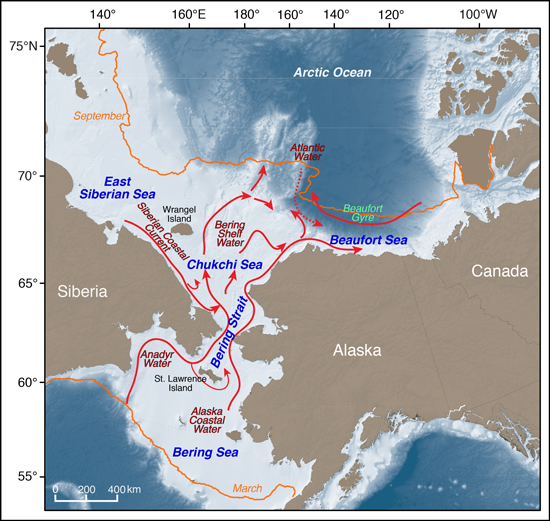
This 5-year effort began with a 3-day workshop to identify science themes and relevant data sets and to form synthesis teams to address high-priority questions about the new normal Pacific Arctic [Sheffield Guy et al., 2012]. More than 100 Arctic scientists and local experts completed the first phase of this effort, an interdisciplinary synthesis of marine ecosystem science in the Pacific Arctic, in August 2015.
The synthesis strengthens scientific understanding of recent, extreme changes in the region’s biophysics, such as sea ice loss and change in occurrence of high-latitude species. It was structured around three themes. The first is observations and models of sea ice loss and effects on primary production—the ability of organisms at the base of the food web to synthesize organic compounds from carbon dioxide. The second is the response of middle trophic species, such as forage fishes, to the new state of the Arctic. Rounding out the synthesis is the third theme: responses of top-level predators—seabirds, seals, walruses, and whales [Moore and Stabeno, 2015].
What Does the New Normal Pacific Arctic Look Like?
Alaskan Arctic winters are expected to remain seasonally icy, but the duration of the summer open-water period will continue to lengthen.
In the future, the Arctic will be a different place than it was even a few decades ago. Alaskan Arctic winters are expected to remain seasonally icy, but the duration of the summer open-water period will continue to lengthen [Wang and Overland, 2015]. Fewer days of sea ice cover and reduced sea ice thickness and extent increase the phytoplankton growing season: Net primary production has increased dramatically, by 30% since the late 1990s across the Arctic Ocean [Arrigo and van Dijken, 2015].
Ocean acidification is also increasing. This will affect commercial and subsistence fisheries because the lower pH makes it difficult for marine organisms to grow and maintain hard shells and skeletons [Mathis et al., 2015].
How Does the New Normal Affect Specific Species?
Middle trophic-level marine communities, including benthic organisms and key forage fishes, have variable responses to the new normal conditions. Benthic “hot spots” identified by high biodiversity and biomass have been tracked over the past 3 decades, with some showing a northward trend in their epicenters [Grebmeier et al., 2015]. Ripple effects from this northward shift extend to all levels of the food web, helping some species and challenging others.

For example, nearshore patches of benthic prey (clams, polychaete worms, crabs), dense enough to be profitable for such predators as eiders, change both spatially and annually, making it difficult to pinpoint key foraging areas to protect [Lovvorn et al., 2015]. Chinook salmon (Oncorhynchus tshawytscha) may be moving northward into the Arctic as conditions become more favorable [Logerwell et al., 2015]. Some populations of ringed and bearded seals (Phoca hispida and Erignathus barbatus, respectively) are growing larger, and their blubber is thicker [Crawford et al., 2015].
Some commercially important fish in the cod family are present in the Arctic but don’t appear to be spawning [Logerwell et al., 2015]. As Arctic cod (Boreogadus saida) follow the receding ice edge, they are moving out of foraging range for an Arctic seabird, the black guillemot (Cepphus grylle), that feeds on them. Black guillemot parents struggle to feed their nestlings, whose health has declined [Divoky et al., 2015]. Average weight of guillemot chicks declined by 25 grams in the new normal (2003–2012) when compared with a historical period (1975–1984).
Top predators (including seabirds, walrus, and whales) are also responding to changing biophysical conditions in the new normal Pacific Arctic. SOAR participants identified seabird and marine mammal hot spots throughout the Pacific Arctic. Seabird and marine mammal hot spots overlapped at the mouth of Barrow Canyon, a submarine canyon off the northern coast of Alaska [Kuletz et al., 2015].
Project participants defined core-use areas, where bowhead whales regularly concentrated, in the Bering-Beaufort-Chukchi seas [Citta et al., 2015]. For the first time, data from acoustic recorders throughout the region were combined to describe the acoustic world encountered by bowheads throughout their annual migration [Clark et al., 2015]. This SOAR study captured recordings of bowheads and bearded seals singing over periods lasting more than a month, the sounds of seismic air guns and high winds, and, of course, quiet periods. A sound sample from the study—the trill of a male bearded seal in the Bering Sea—can be heard below.
Bearded seals vocalized more frequently in areas with more sea ice and during the mating season. Thus, sea ice loss may negatively affect the behavioral ecology of this species [MacIntyre et al., 2015]. Growth and body condition (an indicator of overall health based on measures of body fat, size, and age) for such other species as ringed seals and belugas have also suffered [Harwood et al., 2015]. In contrast, bowhead whale body condition has improved approximately 2.7% per decade since 1989, coincident with sea ice loss, likely due to more favorable conditions for the whales’ zooplankton prey [George et al., 2015].
Beyond the New Normal

Changes in marine bird and mammal habitats and health reflect changes in the ecosystem as a whole. We can look to top predators as sentinels of this change. For example, a shift from a Pacific Arctic driven by benthic production on the ocean’s floor to one dominated by burgeoning pelagic life near the surface is reflected in the abundance and health of these animals. This shift will also attract new species, such as killer whales (Orcinus orca), not historically observed in the region.
Monitoring and predicting conditions in the Pacific Arctic are therefore top priorities. To these ends, a SOAR contribution by Moore and Stabeno [2015] introduced the Arctic Marine Pulses (AMP) conceptual model, which depicts seasonal “pulses” of nutrients, zooplankton, and forage fishes transported from the northern Bering Sea through Bering Strait, across the Chukchi Sea, and into the Beaufort Sea. SOAR participants plan to further refine the AMP model by investigating variability in atmospheric forcing of inflow at Bering Strait and summarizing currently known changes to pelagic-benthic coupling, transport, and nutrient upwelling events in the Chukchi and Beaufort seas.
The second and final phase of SOAR will extend the synthesis effort and culminate in a new special issue of the journal Deep-Sea Research Part II: Topical Studies in Oceanography. Papers for the issue are currently in review, with a total of 16 contributed articles expected. Complete lists of SOAR Phase II special issue papers, contributors, and associated products are available online.
Acknowledgments
The SOAR project is supported by the Bureau of Ocean Energy Management (BOEM) and the National Oceanic and Atmospheric Administration and led by Sue Moore, Phyllis Stabeno, and an 11-member science steering committee.
References
Arrigo, K. R., and G. L. van Dijken (2015), Continued increases in Arctic Ocean primary production, Prog. Oceanogr., 136, 60–70, doi:10.1016/j.pocean.2015.05.002.
Citta, J. J., et al. (2015), Ecological characteristics of core-use areas used by Bering-Chukchi-Beaufort (BCB) bowhead whales, 2006–2012, Prog. Oceanogr., 136, 201–222, doi:10.1016/j.pocean.2014.08.012.
Clark, C. W., C. L. Berchok, S. B. Blackwell, D. E. Hannay, J. Jones, D. Ponirakis, and K. M. Stafford (2015), A year in the acoustic world of bowhead whales in the Bering, Chukchi and Beaufort seas, Prog. Oceanogr., 136, 223–240, doi:10.1016/j.pocean.2015.05.007.
Crawford, J. A., L. T. Quakenbush, and J. J. Citta (2015), A comparison of ringed and bearded seal diet, condition and productivity between historical (1975–1984) and recent (2003–2012) periods in the Alaskan Bering and Chukchi seas, Prog. Oceanogr., 136, 133–150, doi:10.1016/j.pocean.2015.05.011.
Divoky, G. J., P. M. Lukacs, and M. L. Druckenmiller (2015), Effects of recent decreases in Arctic sea ice on an ice-associated marine bird, Prog. Oceanogr., 136, 151–161, doi:10.1016/j.pocean.2015.05.010.
Frey, K. E., G. W. K. Moore, L. W. Cooper, and J. M. Grebmeier (2015), Divergent patterns of recent sea ice cover across the Bering, Chukchi, and Beaufort seas of the Pacific Arctic region, Prog. Oceanogr., 136, 32–49, doi:10.1016/j.pocean.2015.05.009.
George, J. C., M. L. Druckenmiller, K. L. Laidre, R. Suydam, and B. Person (2015), Bowhead whale body condition and links to summer sea ice and upwelling in the Beaufort Sea, Prog. Oceanogr., 136, 250–262, doi:10.1016/j.pocean.2015.05.001.
Grebmeier, J. M., et al. (2015), Ecosystem characteristics and processes facilitating persistent macrobenthic biomass hotspots and associated benthivory in the Pacific Arctic, Prog. Oceanogr., 136, 92–114, doi:10.1016/j.pocean.2015.05.006.
Harwood, L. A., T. G. Smith, J. C. George, S. J. Sandstrom, W. Walkusz, and G. J. Divoky (2015), Change in the Beaufort Sea ecosystem: Diverging trends in body condition and/or production in five marine vertebrate species, Prog. Oceanogr., 136, 263–273, doi:10.1016/j.pocean.2015.05.003.
Kuletz, K. J., M. C. Ferguson, B. Hurley, A. E. Gall, E. A. Labunski, and T. C. Morgan (2015), Seasonal spatial patterns in seabird and marine mammal distribution in the eastern Chukchi and western Beaufort seas: Identifying biologically important pelagic areas, Prog. Oceanogr., 136, 175–200, doi:10.1016/j.pocean.2015.05.012.
Logerwell, E., et al. (2015), Fish communities across a spectrum of habitats in the western Beaufort Sea and Chukchi Sea, Prog. Oceanogr., 136, 115–132, doi:10.1016/j.pocean.2015.05.013.
Lovvorn, J. R., A. R. Rocha, S. C. Jewett, D. Dasher, S. Oppel, and A. N. Powell (2015), Limits to benthic feeding by eiders in a vital Arctic migration corridor due to localized prey and changing sea ice, Prog. Oceanogr., 136, 162–174, doi:10.1016/j.pocean.2015.05.014.
MacIntyre, K. Q., K. M. Stafford, P. B. Conn, K. L. Laidre, and P. L. Boveng (2015), The relationship between sea ice concentration and the spatio-temporal distribution of vocalizing bearded seals (Erignathus barbatus) in the Bering, Chukchi, and Beaufort seas from 2008 to 2011, Prog. Oceanogr., 136, 241–249, doi:10.1016/j.pocean.2015.05.008.
Mathis, J. T., S. R. Cooley, N. Lucey, S. Colt, J. Ekstrom, T. Hurst, C. Hauri, W. Evans, J. N. Cross, and R. A. Feely (2015), Ocean acidification risk assessment for Alaska’s fishery sector, Prog. Oceanogr., 136, 71–91, doi:10.1016/j.pocean.2014.07.001.
Moore, S. E., and P. J. Stabeno (2015), Synthesis of Arctic Research (SOAR) in marine ecosystems of the Pacific Arctic, Prog. Oceanogr., 136, 1–11, doi:10.1016/j.pocean.2015.05.017.
Sheffield Guy, L., S. E. Moore, and P. J. Stabeno (2012), Marine ecosystem synthesis: From physics to whales in the Pacific Arctic, Eos Trans. AGU, 93(48), 501, doi:10.1029/2012EO480007.
Wang, M., and J. E. Overland (2015), Projected future duration of the sea-ice-free season in the Alaskan Arctic, Prog. Oceanogr., 136, 50–59, doi:10.1016/j.pocean.2015.01.001.
Wood, K. R., N. A. Bond, S. L. Danielson, J. E. Overland, S. A. Salo, P. J. Stabeno, and J. Whitefield (2015), A decade of environmental change in the Pacific Arctic region, Prog. Oceanogr., 136, 12–31, doi:10.1016/j.pocean.2015.05.005.
Author Information
Lisa Sheffield Guy, Joint Institute for the Study of the Atmosphere and Ocean, University of Washington, Seattle; Sue E. Moore, National Marine Fisheries Service, National Oceanic and Atmospheric Administration (NOAA), Seattle, Wash.; email: [email protected]; and Phyllis J. Stabeno, Pacific Marine Environmental Laboratory, NOAA, Seattle, Wash.
Citation: Sheffield Guy, L., S. E. Moore, and P. J. Stabeno (2016), What does the Pacific Arctic’s new normal mean for marine life?, Eos, 97, doi:10.1029/2016EO051731. Published on 9 May 2016.
Text © 2016. The authors. CC BY-NC 3.0
Except where otherwise noted, images are subject to copyright. Any reuse without express permission from the copyright owner is prohibited.